Inter la ekzistantaj vivantaj estaĵoj, birdoj kaj mamuloj estas homootermaj (krom nur nudaj molaj ratoj). Krome, la 15an de majo 2015, oni malkovris la unuajn tute varmajn sangajn fiŝojn, kiujn sciencistoj de la Nacia Oceana kaj Atmosfera Administracio de Usono malkovris. La demando pri ĉu pterosaŭroj kaj dinosaŭroj apartenis al varmaj sangaj bestoj estas ankaŭ diskutebla, kvankam lastatempe esploristoj pli kaj pli inklinas al varma sango, kaj debatoj estas jam pri kiuj el la specioj estis varma sango. Ankaŭ ne ekzistas fina klareco pri kia endotermo posedis la dinosaŭroj, sed la disponeblaj datumoj permesas konkludi, ke grandaj dinosaŭroj havis almenaŭ inerian homoyotermion.
Hodiaŭ, plej multaj esploristoj opinias, ke en sia metabola reĝimo, dinosaŭroj okupis ne nur interan pozicion inter bestoj kun "varma sango" kaj "malvarma sango", sed esence diferencis de ambaŭ. Observoj de grandaj modernaj reptilioj montris, ke se besto havas malpliigitan korpan grandon de pli ol 1 m (nome, preskaŭ ĉiuj dinosaŭroj estis tiel), tiam en ebla kaj varma (subtropika) klimato kun malgrandaj ĉiutagaj temperaturaj fluktuoj, ĝi tute kapablas subteni konstantan korpan temperaturon supre 30 ° C: la varmokapacito de akvo (el kiu la korpo konsistas el 85%) estas sufiĉe granda, ke ĝi simple ne havas tempon por malvarmigi dumnokte. La ĉefa afero estas, ke ĉi tiu alta korpa temperaturo estas certigita nur pro varmego el la ekstero, sen ia ajn implikiĝo de sia propra metabolo (por kiu mamuloj devas elspezi 90% de la manĝaĵo, kiun ili konsumas). Do, besto kun grandecoj tipaj de plej multaj dinosaŭroj povas atingi la saman gradon de temperaturregulado kiel mamuloj, konservante tipe reptilian metabolan indicon, ĉi tiu fenomeno J. Hotton (1980) nomis inercia homeotermio. Ŝajne, ĝi estis ĝuste la inercia homoyotermio (kunigita al bipedeco), kiu igis la dinosaŭrojn reĝoj de mesozoika naturo.
En nova studo, kanadaj kaj brazilaj sciencistoj eble trovis indikon pri ĉi tiu evolua mistero. Teamo gvidata de Glenn Tattersall de la Universitato Brock trovis, ke la argentina nigra kaj blanka tagu (Salvator merianae) havas sezonan varman sangon. Ĉi tiu lacerto, ĝis 150 centimetroj longa, loĝas en plej granda parto de Sudameriko kaj estas konata de biologoj. Dum la plej granda parto de la jaro, kiel multaj aliaj reptilioj, la teko ĉirkaŭbrakas sunon dumtage, kaj nokte ili kaŝas sin en truoj kaj malvarmas. Tamen sciencistoj, kiuj uzas sensilojn kaj varmajn ĉambrojn, trovis, ke dum la reprodukta sezono, de septembro ĝis decembro, en la matenaj horoj, la spira ritmo kaj kora ritmo de la besto pliiĝas, kaj ilia temperaturo altiĝas, pli kaj pli ol la temperaturo en la truo je dek gradoj Celsius. Sciencistoj opinias, ke sudamerikaj lacertoj estas intera ligo inter bestoj de malvarma sango kaj varma sango. Pliigo de korpa temperaturo dum la reprodukta sezono pliigas ilian aktivecon serĉinte kompanion, akcelas la disvolviĝon de ovoj kaj permesas al vi pli atenti la idaron. Krome, ekzemple, korŝira testudo, pro la laboro de muskoloj, izolanta grasa tavolo kaj grandaj grandecoj, konservas korpan temperaturon pli alte ol la temperaturo de la ĉirkaŭa akvo. Grandaj kontrolaj lacertoj ankaŭ varmiĝas dum ĉasado aŭ aktiva movado. Grandaj serpentoj kiel pitoj kaj boacoj povas pliigi korpan temperaturon kurbante en ringo kaj kontraktante muskolojn, ĉi tio kutimas varmigi kaj elovi ovojn.
Specoj de hejmotermio
Distingu vera kaj inercia hejmotermo.
- Vera hejmotermo okazas, kiam vivanta estaĵo havas sufiĉan nivelon de metabolo por konservi konstantan korpan temperaturon pro la sendependa produktado de energio de la konsumita manĝaĵo. Modernaj birdoj kaj mamuloj estas veraj hejmotermaj kreitaĵoj. Krom sufiĉaj energifontoj, ili ankaŭ havas diversajn me mechanismsanismojn desegnitajn por reteni varmon (plumoj, lano, subkutana tavolo de adiposa histo) kaj por protekti kontraŭ varmado kontraŭ altaj mediaj temperaturoj (ŝvito). La malavantaĝo de ĉi tiu mekanismo estas, ke necesas multe da energio por konservi korpan temperaturon, kaj tial la bezono de manĝaĵo estas pli alta ol en ĉiu alia kazo.
- Inercia homoyotermio - ĉi tio subtenas konstantan korpan temperaturon pro la granda grandeco kaj granda korpa pezo, same kiel specifan konduton (ekzemple, ŝoviĝu en la suno, malvarmu en akvo). La efikeco de la inercia endotermia mekanismo dependas ĉefe de la proporcio de varmokapacito (simpligita - maso) kaj averaĝa varmofluo tra la korpa surfaco (simpligita - korpa areo), tial ĉi tiu mekanismo povas esti klare observata nur en grandaj specioj. La inercia homoyoterma estaĵo malrapide varmiĝas dum periodoj de temperaturpliiĝo, kaj malrapide malvarmas dum periodoj malvarmetantaj, tio estas, pro la alta varmokapacito, la korpaj temperaturaj fluktuoj malpliiĝas. La malavantaĝo de inercia homoyotermio estas, ke eblas nur kun certa speco de klimato - kiam la meza ĉirkaŭa temperaturo respondas al la dezirata korpa temperaturo kaj ne ekzistas longaj periodoj de severa malvarmigo aŭ varmiĝo. El la avantaĝoj, malgranda bezono de manĝaĵo devas esti emfazita kun sufiĉe alta agnivelo. Karakteriza ekzemplo de inercia homeotermio estas krokodilo. La haŭto de la krokodilo estas kovrita de rektangulaj kornaj ŝildoj, kiuj estas aranĝitaj en regulaj vicoj sur la dorso kaj abdomeno, sub ili en la dorsopovo kaj malpli ofte en la abdomina parto osteodermo, formante karapaceon. Dumtage, osteodermoj amasigas varmon kun sunlumo. Pro tio, la korpa temperaturo de granda krokodilo dum la tago povas flosi ene de nur unu aŭ du gradoj. Kune kun krokodiloj, kondiĉo proksima al inercia homeotermio povas esti observata en la plej grandaj teraj kaj maraj testudoj, same kiel Komodo lacertoj, grandaj pitonoj kaj boacoj.
Homoyotermaj bestoj
Homootermaj bestoj (varmaj sangaj organismoj) estas bestoj, kies temperaturo estas pli-malpli konstanta kaj, kutime, ne dependas de la medio-temperaturo. Ĉi tiuj inkluzivas mamulojn kaj birdojn, en kiuj la konstanteco de temperaturo asocias kun pli alta metabolan indicon kompare kun poikilotermaj organismoj. Krome ili havas termikan izolan tavolon (plumaro, pelto, graso). Ilia temperaturo estas relative alta: ĉe mamuloj ĝi estas 36–37 ° С, kaj ĉe birdoj ripozaj ĝis 40–41 ° С.
POIKILOTERMAJ ANIMALOJ - [ĉ. poikilos motley, diversa + termika varmo, varmego] - malvarmaj sangaj bestoj, bestoj kun nestabila korpa temperaturo, kiu varias depende de la medio-temperaturo, ĉi tiuj inkluzivas ĉiujn senvertebrulojn, kaj ankaŭ fiŝojn, amfibiojn, reptiliojn kaj individuajn mamulojn. )
Dum evoluo, homoyotermaj bestoj disvolvis la kapablon defendi sin de la malvarmo (migrado, hibernado, pelto, ktp.).
Ni jam scias, ke hejmotermaj bestoj povas subteni korpan temperaturon en multe pli vasta temperaturo ol poikilotermaj bestoj (vidu Fig. 3), tamen ambaŭ mortas je proksimume la samaj temperaturoj tre altaj aŭ tro malaltaj (en la unua kazo, pro proteina koaguliĝo, kaj en la dua - pro glaciado de intracelulaj akvoj kun formado de glaciaj kristaloj). Sed ĝis tio okazis, ĝis la temperaturo atingis kritikajn valorojn, la korpo luktas por konservi ĝin normale aŭ almenaŭ proksime al normala nivelo. Nature, ĉi tio estas plene karakteriza por hejmotermaj organismoj kun termoregulado, kapablaj plibonigi aŭ malfortigi ambaŭ varmoproduktadon kaj varmotransporton depende de kondiĉoj. Varmotransigo estas pure fiziologia procezo, ĝi okazas ĉe la organo kaj organismo niveloj, kaj varmoproduktado baziĝas sur fiziologiaj, kemiaj kaj molekulaj mekanismoj. Antaŭ ĉio temas pri malvarmoj, malvarmaj tremoj, t.e., malgrandaj kuntiriĝoj de skeletaj muskoloj kun malalta koeficiento kaj pliigita varmoproduktado. La korpo ŝaltas ĉi tiun mekanismon aŭtomate, reflekte. Ĝia efiko povas esti plibonigita per aktiva libervola muskola agado, kiu ankaŭ plibonigas varmegan generacion. Estas ne hazarde, ke por varmiĝi, ni recidas al movado.
Korpa temperaturo. Homotermaj bestoj ne nur provizas varmon pro sia propra varmoproduktado, sed ankaŭ kapablas aktive reguligi ĝian produktadon kaj konsumon. Pro tio, ili karakterizas sin per alta kaj sufiĉe stabila korpa temperaturo. Ĉe birdoj, la plej profunda korpa temperaturo estas kutime ĉirkaŭ 41 ° C kun fluktuoj en malsamaj specioj de 38 ĝis 43.5 ° C (datumoj por 400 vvd). Sub kondiĉoj de kompleta ripozo (ĉefa metabolo), ĉi tiuj diferencoj estas iom mildigitaj, kiuj iras de 39,5 ĝis 43,0 ° С. Je la nivelo de individua organismo, korpa temperaturo montras altan stabilecon: la gamo de ĝiaj ĉiutagaj ŝanĝoj kutime ne superas 2-4 ° C, kaj ĉi tiu fluktuado ne rilatas al aera temperaturo, sed reflektas la rtm de metabolo. Eĉ en arktaj kaj antarktaj specioj, ĉe ĉirkaŭaj temperaturoj ĝis 20–50 ° C, korpa temperaturo varias inter la samaj 2–4 ° C.
Adaptiĝaj procezoj en bestoj koncerne temperaturon kaŭzis aperon de poikilotermaj kaj homoyotermaj bestoj. La superforta plimulto de bestoj estas lotermikoj, tio estas, ke la temperaturo de siaj propraj korpoj ŝanĝiĝas kun la temperaturo de la medio: amfibioj, reptilioj, insektoj, ktp. Multe pli malgranda proporcio de bestoj estas homoyotermaj, tio estas, ili havas konstantan korpan temperaturon, sendepende de temperaturo. ekstera medio: mamuloj (inkluzive homoj) havantaj korpan temperaturon de 36–37 ° С, kaj birdoj kun korpa temperaturo de 40 ° С.
| Fiziologia adapto de homeoterma besto al malvarmo. | 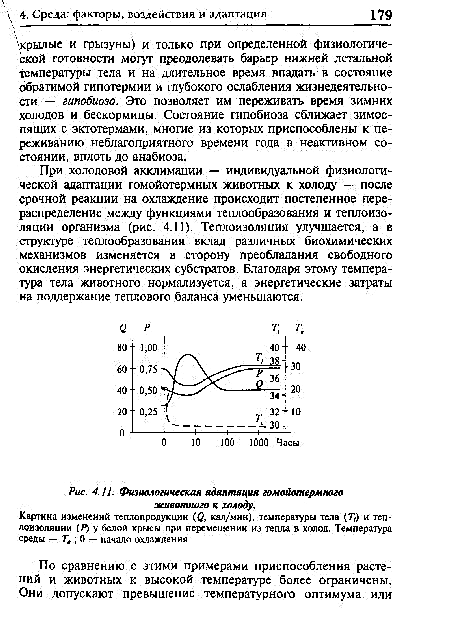 |
Sed nur veraj sangaj hejmaj bestoj - birdoj kaj mamuloj povas subteni konstantan altan korpan temperaturon kun signifaj ŝanĝoj de la temperaturo. Ili havas perfektajn nervajn kaj hormonajn mekanismojn de aktiva varmiga regulado, kiuj inkluzivas ne nur rimedojn por efika reguligo de varmotransportado (per ŝanĝoj en ekstercentra sanfluo, spirado, ŝvitado kaj varmokondukado de la haroj), sed ankaŭ ŝanĝoj de intenseco de oksidaj procezoj kaj varmoproduktado en la korpo. Pro tio, la temperaturo de la internaj partoj de la korpo ne multe dependas de la temperaturo de la medio. Tial, birdoj kaj mamuloj ankaŭ nomiĝas endotermaj organismoj. En iuj el ili, termoregulaj mekanismoj atingas grandan potencon. Do, polusa vulpo, polusa strigo kaj blanka ansero facile toleras severan malvarmon sen falo en korpa temperaturo kaj konservante diferencon en 100 kaj pli da temperaturoj de korpo kaj medio. Pro la dikeco de subkutana graso kaj la proprecoj de ekstercentra sangocirkulado, multaj pinipiedoj kaj balenoj estas bonege adaptitaj por longa restado en glacia akvo.
Do, adaptaj ŝanĝoj en varmotransportado en hejmotermaj bestoj povas celi ne nur konservi altan nivelon de metabolo, kiel en plej multaj birdoj kaj mamuloj, sed ankaŭ fiksi malaltan nivelon de metabolo en kondiĉoj, kiuj minacas malpliiĝon de energiaj rezervoj. Ĉi tiu kapablo ŝanĝi la tipojn de reguligo de varmotransigo signife vastigas la ekologiajn eblecojn bazitajn sur homaotermio.
Aktiva vivo ĉe temperaturoj sub nulo povas konduki nur homotermajn bestojn. Poikilotermaj kvankam ili eltenas temperaturojn signife sub nulo, sed samtempe perdas sian moveblecon. Temperaturo de la ordo de +40 ° C, t.e. eĉ pli malalta ol la koagula temperaturo de la proteino, estas ekstrema por la plej multaj bestoj.
Kaze de Malvarma Aŭlimigo - individua fiziologia adapto de hejmotermaj bestoj al malvarmo - post urĝa reago al malvarmigo, laŭgrada redistribuo okazas inter la funkcioj de varmega generado kaj termika izolado de la korpo (Fig. 4.11). Termika izolado plibonigas, kaj en la strukturo de varmo-generacio, la kontribuo de diversaj biokemiaj mekanismoj ŝanĝiĝas en la direkto al la prevalenco de libera oksido de energiaj substratoj. Pro tio, la korpa temperaturo de la besto estas normaligita, kaj la energiaj kostoj por konservi la varman ekvilibron.
Esence malsama speco de adaptiĝo al la temperatura faktoro estas karakteriza por homoteraj bestoj. Iliaj temperaturaj adaptoj estas asociitaj kun aktiva bontenado de konstanta interna temperaturo kaj baziĝas sur alta nivelo de metabolo kaj efika reguliga funkcio de la centra nerva sistemo. La komplekso de morfofiziologiaj mekanismoj por konservi la termikan homeostazon de la korpo estas specifa propraĵo de hejmotermaj bestoj.
Se poikilotermoj estas entombigaj, tiam vintra kaj somera hibernado estas eneca en homoyotermaj bestoj, kies fiziologiaj kaj molekulaj mekanismoj diferencas de entumeco. Iliaj eksteraj manifestiĝoj estas samaj: malpliiĝo de korpa temperaturo preskaŭ al ĉirkaŭa temperaturo (nur dum vintra hibernado, dum somera hibernado ĝi ne) kaj metabola indico (10-15 fojojn), ŝanĝiĝo de la reago de la interna medio de la korpo al alkala flanko, malkresko de la ekscitebleco de la spira centro kaj malpliigo de la spirado al 1-inspiro en 2,5 minutoj, la ritmo cardiaco ankaŭ falas akre (ekzemple, en vespertoj de 420 ĝis 16 korbatoj / min). La kialo de tio estas pliigo de la tono de la parasimpatia nerva sistemo kaj malkresko de la simpatia ekscitebleco. La plej grava afero estas, ke dum hibernado la termoregula sistemo estas malŝaltita. La kialoj de ĉi tio estas malpliigo de la agado de tiroida glando kaj malpliigo de la enhavo de tiroidesaj hormonoj en la sango. Homoyotermaj bestoj fariĝas poikilotermaj.
Birdoj kaj mamuloj kapablas konservi sufiĉe konstantan korpan temperaturon, sendepende de la medio. Ĉi tiuj bestoj estas nomataj homokotermaj (el la greka. Homoyotermaj bestoj relative malmulte dependas de eksteraj varmofontoj. Pro la alta interŝanĝo ili generas sufiĉan varmon, kiu povas stoki. Ĉar ĉi tiuj bestoj ekzistas pro internaj varmofontoj, ili ofte ofte nomiĝas endotermaj). .
Ĉio ĉi-supre rilatas al la nomata profunda korpa temperaturo, kiu karakterizas la termikan staton de la termostatike kontrolita "kerno" de la korpo. Ĉe ĉiuj homotermaj bestoj, la eksteraj tavoloj de la korpo (integro, parto de la muskoloj, ktp.) Formas pli-malpli prononcan "ŝelon", kies temperaturo vaste varias. Tiel stabila temperaturo karakterizas nur la areon de lokalizado de gravaj internaj organoj kaj procezoj. Surfacaj histoj rezistas pli akuratajn temperaturojn.La egoo povas esti utila por la korpo, ĉar en tia situacio la temperaturo-gradiento ĉe la limo inter la korpo kaj la medio malpliiĝas, kio ebligas konservi termikan homeostazon de la "kerno" de la korpo kun malpli altaj energiaj elspezoj.
La liberigo de energio en formo de varmo akompanas la funkcian ŝarĝon de ĉiuj organoj kaj histoj (Tabelo 4.2) kaj estas karakteriza por ĉiuj vivantaj organismoj. La specifaĵo de hejmotermaj bestoj estas, ke ŝanĝo de varmoproduktado kiel reago al ŝanĝiĝanta temperaturo reprezentas en ili specialan reagon de la korpo, kiu ne influas la nivelon de funkciado de la bazaj fiziologiaj sistemoj.
LANDSCAPA HOMEOSTASIS Kapableco de pejzaĝo reteni en siaj bazaj ecoj sian strukturon kaj la naturon de la rilatoj inter elementoj malgraŭ eksteraj influoj. Hejm-Termaj Animoj [de ĉ. Iotoyuz estas similaj, identaj kaj (Yeghts - varmego], varmaj sangaj bestoj - bestoj, kies korpotemperaturo estas konstante konstante sendepende de la medio-temperaturo pro la energio liberigita dum la metabolo (birdoj kaj mamuloj).
La efiko de medio-temperaturo. Esenca en la disvolviĝo kaj esenca agado de histoj, organoj kaj la korpo entute estas la konstanteco de korpa temperaturo, (homootermaj) bestoj. Homootermaj bestoj distingiĝas per la evolue disvolvita kapablo ŝanĝi la kvanton de varmotransportado (fizika termoregulado) per regulado de sango-cirkulado en surfacaj histoj kaj elvaporiĝo de humido de la korpo, same kiel ŝanĝi varman generacion (kemia termoregulado) konservante konstantan temperaturon de la histoj kaj de la tuta korpo. La relativa konstanteco de la korpa temperaturo de hejmaj bestoj estas subtenata de kompleksa, neŭromumorala regulado de la procezoj de varmogenerado kaj varmo-translokigo. Kiam la korpo malvarmas en la korpo, metabolaj procezoj intensiĝas kaj varmo-generacio pliiĝas, kaj varmotransdono malpliiĝas, kiam varmite kontraŭe, varmoproduktado malpliiĝas kaj varmo-translokigo pliiĝas.
Specioj-diferencoj en la temperatura sojlo preter kiu la normala funkciado de la spermovada aparato estas ĝenata, precipe prononcata kiam oni komparas spermon kun poikilotermaj kaj homoyotermaj bestoj, klarigeblas alimaniere (Holwill, 1969). Unue, malsamaj organismoj povas havi variaĵojn en la strukturo de la enzimo, la nombro kaj speco de ligoj damaĝitaj de termika denaturado de ĝiaj molekuloj. Due, la enzimo en la studataj bestaj specioj povas esti identa, kaj diferencoj en temperaturlimoj ĉe kiuj oni observas lian denaturadon probable estas pro la malsimileco de mediaj kondiĉoj (pH, jona koncentriĝo, ktp.).
Aero kiel vivmedio havas certajn ecojn: tio gvidas la ĝeneralajn evoluajn vojojn de la loĝantoj de ĉi tiu medio. Tiel alta enhavo de oksigeno (ĉirkaŭ 21% en atmosfera aero, iomete malpli en la aero pleniganta la spira sistemo de bestoj) determinas la eblecon formi altan energian metabolon. Estas ne hazarde, ke en ĉi tiu medio ekestis homootermaj bestoj, karakterizitaj de alta energio de la korpo, alta grado de aŭtonomio de eksteraj influoj kaj alta biologia aktiveco en ekosistemoj. Aliflanke, atmosfera aero karakterizas per malalta kaj ŝanĝiĝema humideco. Ĉi tiu cirkonstanco plejparte limigis la eblecojn disvolvi la aeran medion, kaj inter la loĝantoj ĝi estis gvidata de la evoluo de la fundamentaj ecoj de la akva-sala metabola sistemo kaj la strukturo de la spira sistemo.
La dua grava avantaĝo por la loĝantoj de vivantaj organismoj estas ilia protekto kontraŭ la rekta efiko de mediaj faktoroj. Interne de la gastiganto, ili praktike ne renkontas la danĝeron sekiĝi, akrajn fluktuojn de temperaturo, signifajn ŝanĝojn de salo kaj osmotikaj reĝimoj, ktp. Tiel en kondiĉoj aparte stabilaj estas internaj loĝantoj de homoyotermaj bestoj. Fluaĵoj en mediaj kondiĉoj influas internajn parazitojn kaj simbolojn nur nerekte, tra la gastiganta organismo.
Homo kiel specio, esence diferenca de ĉiuj antaŭaj specioj, ekestis en la procezo de evoluo sub la influo de leĝoj komunaj al ĉiuj vivantaj estaĵoj rezulte de fundamenta genetike fiksita malkovro en la procezo de evoluo de la organismoj de la biosfero. Tiaj kardinalaj eltrovaĵoj, kaŭzantaj la aperon de esence novaj specioj, okazis antaŭ la apero de homo. Do, estis plurĉelaj organismoj, vertebruloj, hejmotermaj bestoj kun konstanta korpa temperaturo.
La listigitaj ekzemploj for de elĉerpi ĉiujn formojn de adapta konduto. Ĉi tio inkluzivas la kapablon de multaj birdoj kaj mamuloj aktive konstrui nestojn, truojn kaj aliajn rifuĝejojn kun favora mikroklimato, la uzon de pozoj, kiuj ŝparas energian konsumon, laŭsezonajn movadojn, la adaptiĝan naturon de ĉiutaga agado, ktp. La tuta komplekso de adaptaj kondutaj reagoj, reduktante la intenson de energia interŝanĝo, vastigas la ekologiajn kapablojn de hejmotermaj bestoj.
La asimilita energio, malpli la energio entenata en elĵetita eltiraĵo de la korpo (feĉoj, urino, ktp), estas la metaboligita energio. Parto de ĝi estas asignita en la formo de tesha en la procezo de digesta manĝaĵo kaj estas aŭ dissemita aŭ uzata por termoregulado. La restanta energio estas dividita en la energion de ekzisto, kiu estas tuj konsumita de la plej oftaj vivoformoj (esence ĝi estas ankaŭ "elspezo por spirado"), kaj la produktema energio, kiu akumuliĝas (almenaŭ provizore) en la amaso de kreskantaj histoj, energiaj rezervoj kaj seksaj produktoj (rizo. . 3.1). La energio de ekzisto konsistas el la kostoj de fundamentaj vivprocezoj (baza metabolo, aŭ baza metabolo) kaj la energio elspezita pri diversaj agadaj formoj. Ĉe homootermaj bestoj, la energia elspezo pri termoregulado aldoniĝas al ĉi tio. Ĉiuj ĉi tiuj energiaj kostoj finiĝas per la disipado de energio en formo de varmego - denove, ĉar ne unu funkcio funkcias kun efikeco de 100%. La energio amasigita en la histoj de la heterotrofo konsistigas la malĉefan produktadon de la ekosistemo, kiu povas esti uzata kiel nutraĵo de konsumantoj de pli altaj mendoj.
La avantaĝoj de hejmotermio
Varm-sangaj bestoj, kutime, ne falas en hibernacion, krom kelkaj esceptoj, kaj ili povas esti aktivaj dum la tuta jaro, manĝante, movante sin kaj protektante sin de predantoj.
Kvankam varmaj sangaj bestoj devas konsumi multan manĝaĵon por resti aktivaj, ili havas energion kaj la rimedojn regi en ĉiuj naturaj lokoj, eĉ en malvarma Antarkto aŭ altaj montaroj. Ili ankaŭ povas vojaĝi pli rapide kaj pli longajn distancojn ol malvarm-sangaj bestoj.
Malavantaĝoj de hejmotermio
Ĉar la korpa temperaturo ĉe varmaj sangaj bestoj restas stabila, ili estas idealaj gastigantoj por multaj parazitoj, kiel vermoj, aŭ mikroorganismoj, inkluzive de bakterioj kaj virusoj, multaj el kiuj povas kaŭzi mortigajn malsanojn.
Ĉar hejmotermaj bestoj liberigas sian propran varmon, grava faktoro estas la rilatumo de maso al korpa surfaco. Granda korpa maso produktas pli da varmego, kaj granda korpa surfaco estas uzata por malvarmetiĝi somere aŭ en pli varma vivmedio, ekzemple la grandegaj oreloj de elefantoj. Tial varmaj sangaj bestoj ne povas esti tiel malgrandaj kiel malvarmaj sangaj insektoj.